 投稿系统
投稿系统

Intercellular transfer of SerpinE2 activates PI3K-AKT and β-catenin signaling to promote cardiac hypertrophy
doi: 10.1515/fzm-2025-0021
-
Abstract:
Background Effective inhibition of pathological cardiac hypertrophy is critical for managing various cardiovascular diseases, especially in cold environments. The communication between cardiomyocytes and fibroblasts, mediated by secreted proteins, plays a significant role in the development and progression of pathological cardiac hypertrophy. Serpin Family E Member 2 (serpinE2), secreted by fibroblasts into the extracellular space, has been implicated in this process. However, whether serpinE2 can be internalized by cardiomyocytes and whether cold exposure influences this process remains unclear. Materials and methods Mice were subjected to cold exposure (4 ℃, 12 h/day for 8 weeks), and cardiac hypertrophy was induced by transverse aortic constriction (TAC). SerpinE2 expression was silenced by short interfering RNA (siRNA). Cardiac fibroblasts were stimulated with angiotensin Ⅱ (Ang Ⅱ) to induce serpinE2 secretion. Exogenous recombinant serpinE2, labeled with DyLight 488 or His-tag, was used to evaluate its internalization and functional role in cardiomyocytes. Internalization was inhibited by using antibodies against serpinE2, heparin, or endocytosis inhibitors (β-cyclodextrin, nystatin, dynasore, and chlorpromazine). Chromatin immunoprecipitation followed by quantitative polymerase chain reaction (PCR) was used to assess the binding of the transcription factor CDX1 to the serpinE2 promoter. Results Cold exposure significantly increased serpinE2 mRNA and protein expression in mouse hearts. SerpinE2 levels were also upregulated in plasma and cardiac tissue following TAC. Knockdown of serpinE2 attenuated TAC-induced hypertrophy, restored left ventricular function, and reduced atrial natriuretic peptide, brain natriuretic peptide, and β-myosin heavy chain fragment levels. Exogenous serpinE2 promoted cardiomyocyte hypertrophy, an effect that was reversed by serpinE2 knockdown. Co-culture with conditioned medium from Ang Ⅱ-stimulated fibroblasts increased serpinE2 expression in cardiomyocytes. Exogenous serpinE2 was internalized via endocytosis, which was inhibited by antibodies, heparin, and endocytosis blockers. Internalized serpinE2 activated the protein kinase B (AKT)/β-catenin pathway in cardiomyocytes. CDX1 bound to the serpinE2 promoter and promoted its transcription in fibroblasts. CDX1 overexpression increased serpinE2 and collagen expression, while its suppression had the opposite effect. Administration of exogenous fibroblast growth factor 4 (FGF4) or overexpression of FGF4 plasmid upregulated CDX1, serpinE2, and collagen expression in fibroblasts. Conclusions SerpinE2 expression is responsive to cold stress and mediates intercellular communication between fibroblasts and cardiomyocytes. Fibroblast-secreted serpinE2 is internalized by cardiomyocytes via endocytosis, promoting hypertrophy through activation of the phosphatidylinositol 3-kinase (PI3K)-AKT/β-catenin pathway. The FGF4-CDX1 axis regulates serpinE2 expression and secretion in cardiac fibroblasts. -
Key words:
- cold exposure /
- serpinE2 /
- cardiac hypertrophy /
- cellular communication /
- endocytosis
-

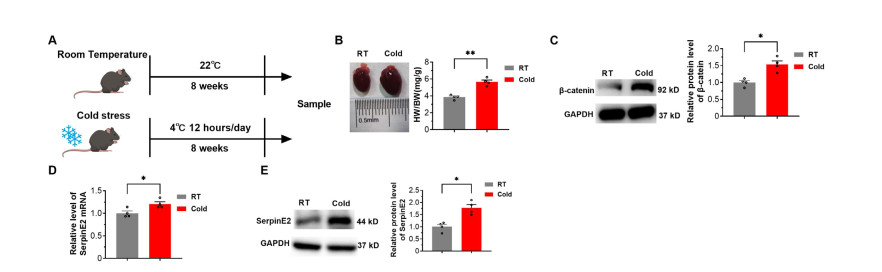
Figure 1. SerpinE2 expression is upregulated during chronic cold exposure
(A) Schematic diagram illustrating the construction of the mouse mode of cold stimulation. (B) Representative image of heart sizes and statistical results of heart weight/body weight ratios (N = 4). (C) Relative protein levels of β-catenin (N = 4). (D) Relative mRNA levels of serpinE2 detected using real-time polymerase chain reaction (PCR) (N = 4). (E): Relative protein levels of serpinE2 in hearts detected by western blot analysis (N = 4). Data are presented as mean ± SEM. *P < 0.05. RT, room temperature. Cold: cold stress (4 ℃).

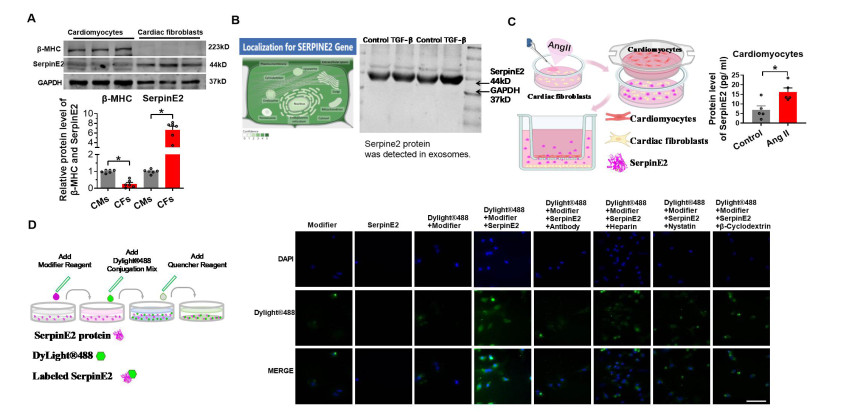
Figure 2. SerpinE2 secreted by fibroblasts is internalized into cardiomyocytes
(A) SerpinE2 and β-myosin heavy chain (MHC) protein levels in cardiomyocytes and cardiac fibroblasts determined using western blot under normal culture conditions (N = 6). (B) Subcellular locations of serpinE2 obtained from the GENECARDs database (
https://www.genecards.org/ ). Exosomes from the supernatant of fibroblasts were identified using Western blot analysis. (C) Cardiac fibroblasts treated with Ang Ⅱ for 24 h and co-cultured with cardiomyocytes for another 24 h. SerpinE2 protein levels in cardiomyocytes measured using an enzyme-linked immunosorbent assay (ELISA) (N = 5). (D) Primary cultured rat cardiomyocytes treated with serpinE2 for 24 h, followed by incubation with serpinE2-antibody, heparin, β-cyclodextrin, and nystatin for 24 h. Results were examined under a fluorescence microscope (Scale bar 50 μm). Data are presented as mean ± SEM. *P < 0.05.
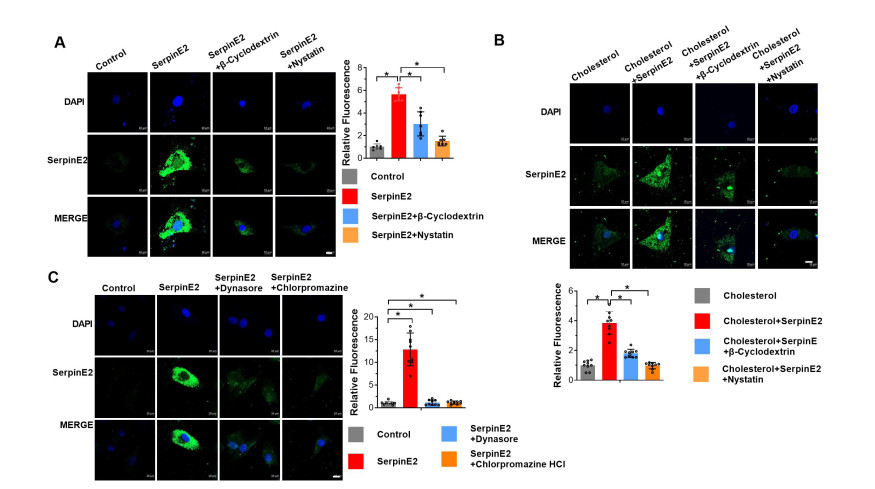
Figure 3. SerpinE2 endocytosis is regulated by both clathrin-dependent and lipid-raft-dependent endocytic mechanisms
(A-B) Primary cultured rat cardiomyocytes treated with serpinE2, serpinE2 + β-cyclodextrin, or serpinE2 + nystatin for 24 h, with or without cholesterol (Scale bar 10 μm, N = 7-9). (C) Primary cultured rat cardiomyocytes treated with serpinE2, serpinE2 + dynasore, or serpinE2 + chlorpromazine for 24 h (Scale bar 10 μm, N = 9). Quantitative results of serpinE2 fluorescence density determined by Image J software v1.0. Data are presented as mean ± SEM. *P < 0.05.

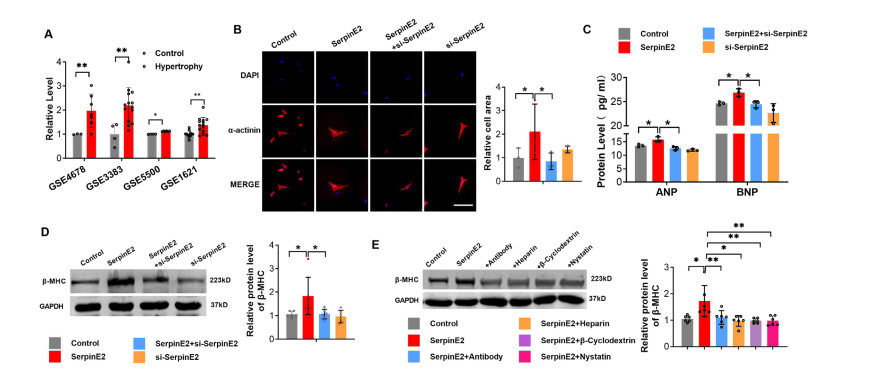
Figure 4. SerpinE2 induces rat cardiomyocyte hypertrophy in vitro
(A) SerpinE2 mRNA levels in cardiac hypertrophy obtained from four mRNA microarray GEO databases (N = 3-14). (B) Primary cultured rat cardiomyocytes treated with serpinE2, serpinE2 + si-serpinE2, or si-serpinE2 for 24 h, and cell cross-sectional area determined using immunofluorescence assay (Scale bar 50 μm, N = 3); (C) Protein levels of ANP and BNP measured using ELISA (N = 3); (D) Relative protein levels of β-myosin heavy chain (β-MHC) determined using western blot analysis (N = 6). (E) Primary cultured rat cardiomyocytes treated with serpinE2, serpinE2 + antibody, serpinE2 + heparin, serpinE2 + β-cyclodextrin, or serpinE2 + nystatin for 24 h. Then the relative protein levels of β-MHC were detected using western blot (N = 6). Data are presented as mean ± SEM. *P < 0.05, **P < 0.01.

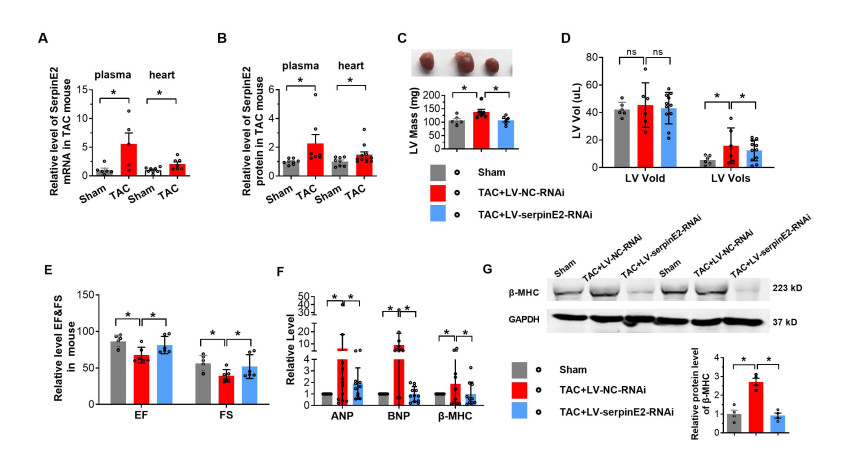
Figure 5. Inhibition of SerpinE2 alleviates pressure overload-induced cardiac hypertrophy
(A-B) The cardiac hypertrophy mouse model was developed using transverse aortic constriction (TAC) methods and relative serpinE2 mRNA levels (N = 5-8) (A) and protein activities (B) in plasma and hearts were determined using real-time polymerase chain reaction (PCR) and ELISA, respectively (N = 7-11). (C) Two weeks after the injection of LV-NC or LV-RNAi-SerpinE2, TAC surgery was performed on mice to induce an in vivo cardiac hypertrophy model for 4 weeks. Representative images of the ventral side and LV mass of the heart (N = 6). (D) Assessments of echocardiographic parameters of end-diastolic volume (Vold) and end-systolic volume (Vols) (N = 6-11). (E)Ejection fraction (EF) and fraction shortening (FS) (N = 6). (F) Relative mRNA levels of ANP, BNP, and β-MHC were detected by real-time PCR (N = 10-11). (G) Relative protein levels of β-MHC in hearts were detected by western blot (N = 4). Data are presented as mean ± SEM. *P < 0.05.

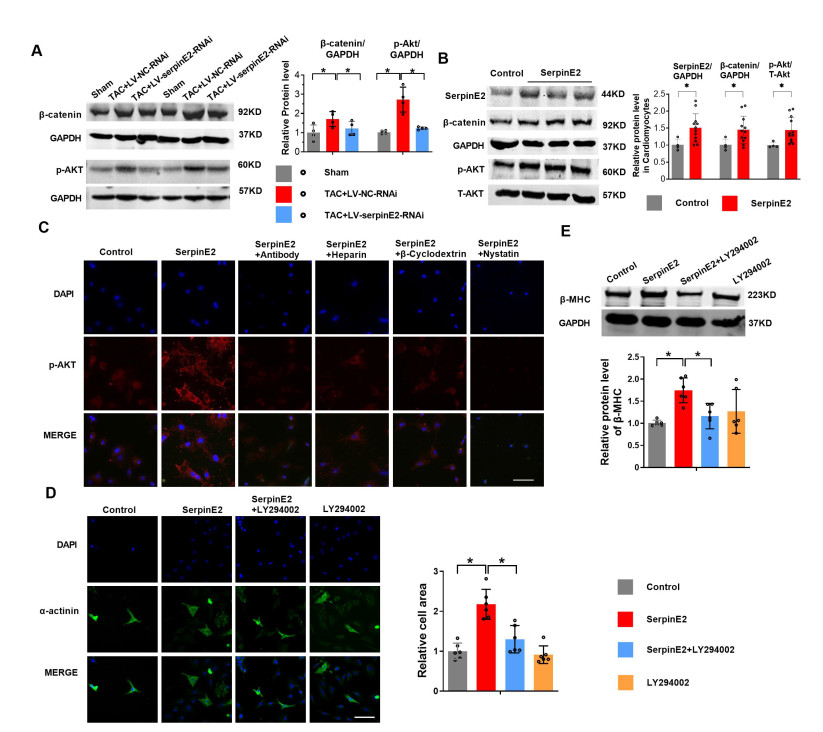
Figure 6. SerpinE2 promotes cardiac hypertrophy by activating the β-catenin and protein kinase B (AKT) pathway
(A) Two weeks after the injection of LV-NC or LV-RNAi-SerpinE2, TAC surgery was performed for 4 weeks to establish a mouse model of cardiac hypertrophy. Relative protein levels of β-catenin, p-AKT detected by western blot (N = 4). (B) Primary cultured rat cardiomyocytes treated with serpinE2 for 24 h. Relative protein levels of serpinE2, β-catenin, p-AKT, and T-AKT detected by western blot (N = 4-12). (C) Primary cultured rat cardiomyocytes treated with serpinE2, serpinE2 + antibody, serpinE2 + heparin, serpinE2 + β-cyclodextrin, or serpinE2 + nystatin for 24 h, and cellular serpinE2 detected by immunofluorescence assay (Scale bar 50 μm). (D) Cardiomyocyte area measured using α-actinin immunofluorescence assay, following the administration of SerpinE2, SerpinE2 + LY294002, or LY294002 for 24 h (N = 6, Scale bar 50 μm). (E) β-MHC detected by western blot, following the administration of SerpinE2, SerpinE2 + LY294002, or LY294002 for 24 h (N = 6). Data are presented as mean ± SEM. *P < 0.05.

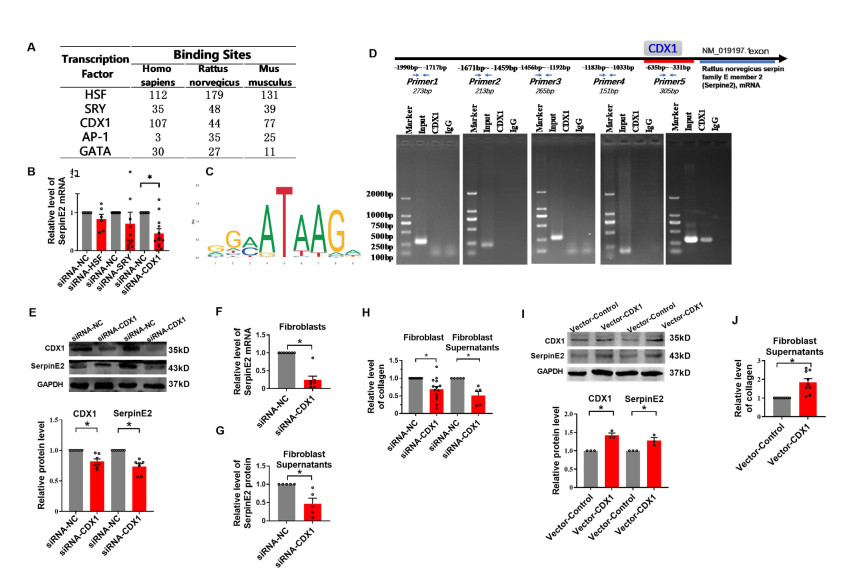
Figure 7. CDX1 upregulates serpinE2 and collagen in rat cardiac fibroblasts
(A) The predicted transcript factors of SerpinE2. (B) SerpinE2 mRNA level in fibroblasts measured by real-time PCR (N = 7). (C) The predicted binding site of CDX1 on the genome of SerpinE2. (D) Schematic diagram of binding sites of CDX1 and primers used for CHIP-PCR using rat SerpinE2 gene sequences and agarose gel electrophoresis from ChIP-PCR showing CDX1 bound to the SerpinE2 promoter in rat cardiac fibroblasts. The promoter regions of SerpinE2 (-1990bp to-331bp) were amplified using the input and immunoprecipitated DNA as templates. (E) Protein levels of SerpinE2 and CDX1 (N = 7). (F) The mRNA level of SerpinE2 after inhibition of CDX1 in fibroblast (N = 7). (G) Protein levels of SerpinE2 in fibroblast supernatants detected using ELISA after inhibition of CDX1 (N = 5). (H) After inhibition of CDX1, collagen contents in fibroblasts or supernatants were measured by Sircol collagen assay (N = 5-14). (I) Protein levels of SerpinE2 and CDX1 in rat cardiac fibroblasts. (J) Collagen contents after overexpression of CDX1 (N = 9). Data are presented as mean ± SEM. *P < 0.05.

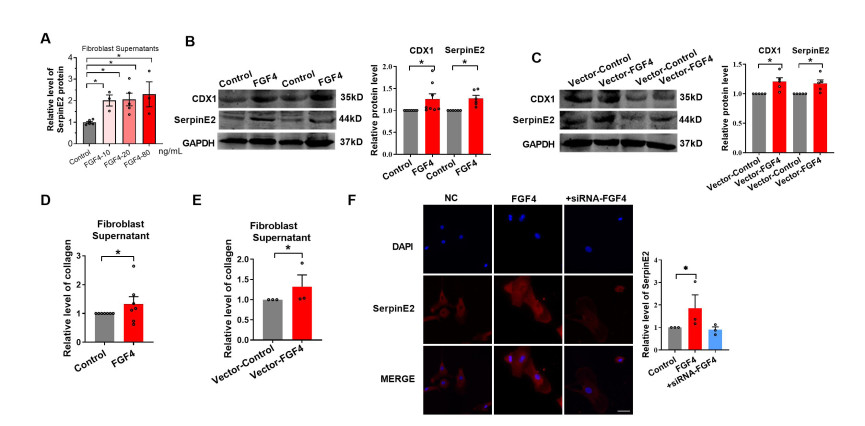
Figure 8. FGF4 upregulates CDX1, serpinE2 and collagen expression in cardiac fibroblasts
(A) SerpinE2 in fibroblast supernatants were detected using an Enzyme-linked Immunosorbent Assay (ELISA) following the administration of 10, 20, or 80 ng/mL FGF4 (N = 3-6). (B-C) Relative protein levels of SerpinE2 and CDX1 in fibroblasts detected by western blot following the treatment with FGF4 (N = 6-8) (B) or FGF4 plasmid transfection (N = 5) (C). (D-E) Collagen contents in fibroblast supernatants determined by Sircol collagen assay following the treatment with FGF4 (N = 7) (D) or FGF4 plasmid transfection (N = 3) (E). (F) SerpinE2 expression in fibroblasts examined using immunofluorescence assay following the treatment with FGF4 or FGF4 + siRNA-FGF (Scale bar 50 μm, N = 3). Data are presented as mean ± SEM. *P < 0.05.

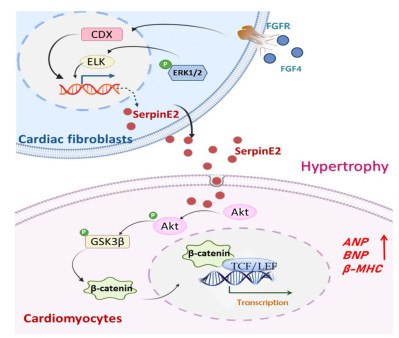
Figure 9. Schematic diagram illustrating the proposed hypothesis on the mechanism of serpinE2 action
; the figure was drawn by Power Point 2021 (Microsoft, Redmond, Washington) FGF4-CDX1 signaling promotes serpinE2 secretion from cardiac fibroblasts. Upon endocytosed into cardiomyocytes, serpinE2 promotes cardiomyocyte hypertrophy by activating the PI3K-AKT/β-catenin pathway; the figure was drawn by Power Point 2021 (Microsoft, Redmond, Washington).
Table 1. Primers used for the Real-Time PCR (qRT-PCR) analysis
RNA name Primers from 5' to 3' ANP-F ACCTGCTAGACCACCTGGAG ANP-R CCTTGGCTGTTATCTTCGGTACCGG BNP-F GAGGTCACTCCTATCCTCTGG BNP-R GCCATTTCCTCCGACTTTTCTC β-MHC-F CCGAGTCCCAGGTCAACAA β-MHC-R CTTCACGGGCACCCTTGGA GAPDH-F GGGGCTCTCTGCTCCTCCCTG GAPDH-R CGGCCAAATCCGTTCACACCG FGF4-F GACCAAGAAGGGGAACCGAG FGF4-R CGGAGGGTCACAGTCTAGGA Cdx1-F TCTACACAGACCACCAACGC Cdx1-R TTTACCTGCCGCTCTGTGAG SerpinE2-F CGATCTCCCTCCCGGTTTC SerpinE2-R TCCCTGTGTTGGAGCCTAGT  下载: 导出CSV
下载: 导出CSV
Table 2. Primers used for the Reverse Transcription PCR (RT-PCR) analysis
RNA name Primers from 5' to 3' CDX1-ChIPF1 GATCATTGGAAACACTGATG CDX1-ChIPR1 GAAAATCTGAGAGCTGCTTTCC CDX1-ChIPF2 TGGAGGCGTGCTCATTTCTGG CDX1-ChIPR2 CAGGGCATCCCTGTGTCCAAAG CDX1-ChIPF3 CTGTAGTCTCCATCTCTGTCTC CDX1-ChIPR3 CCCTCTCAGGTCACAGTCAGAC CDX1-ChIPF4 CAGGAGCATTTGGGACAGAGGG CDX1-ChIPR4 GTAGGGACAAATGCTATGGAAG CDX1-ChIPF5 GTATAGGTTCAAAACACCTGG CDX1-ChIPR5 GGAGTGGGCGACACCGAAGCC negative control-F TTCCCTCAGAACAATAACGCAG negative control-R CCTTCCAAGTAGAAGCTTGGAATG
下载: 导出CSV
-
[1] Shor E, Roelfs D. Climate shock: Moving to colder climates and immigrant mortality. Soc Sci Med, 2019; 235: 112397. doi: 10.1016/j.socscimed.2019.112397 [2] Moghadamnia M T, Ardalan A, Mesdaghinia A, et al. Ambient temperature and cardiovascular mortality: a systematic review and meta-analysis. PeerJ, 2017; 5: e3574. doi: 10.7717/peerj.3574 [3] Fan J F, Xiao Y C, Feng Y F, et al. A systematic review and meta-analysis of cold exposure and cardiovascular disease outcomes. Front Cardiovasc Med, 2023; 10: 1084611. doi: 10.3389/fcvm.2023.1084611 [4] Heidenreich P A, Bozkurt B, Aguilar D, et al. 2022 AHA/ACC/HFSA guideline for the management of heart failure: a report of the American College of Cardiology/American Heart Association Joint Committee on Clinical Practice Guidelines. Circulation, 2022; 146(13): e185. [5] Saucerman J J, Tan P M, Buchholz K S, et al. Mechanical regulation of gene expression in cardiac myocytes and fibroblasts. Nat Rev Cardiol, 2019; 16(6): 361-378. doi: 10.1038/s41569-019-0155-8 [6] Kakkar R, Lee R T. The IL-33/ST2 pathway: therapeutic target and novel biomarker. Nat Rev Drug Discov, 2008; 7(10): 827-840. doi: 10.1038/nrd2660 [7] Zhang X, Sun S, Ren G, et al. Advances in intercellular communication mediated by exosomal ncRNAs in cardiovascular disease. Int J Mol Sci, 2023; 24(22): 16197. doi: 10.3390/ijms242216197 [8] Bouton M C, Boulaftali Y, Richard B, et al. Emerging role of serpinE2/protease nexin-1 in hemostasis and vascular biology. Blood, 2012; 119(11): 2452-2457. doi: 10.1182/blood-2011-10-387464 [9] Zhang J, Luo A, Huang F, et al. SERPINE2 promotes esophageal squamous cell carcinoma metastasis by activating BMP4. Cancer Lett, 2020; 469: 390-398. doi: 10.1016/j.canlet.2019.11.011 [10] Öner M G, Rokavec M, Kaller M, et al. Combined inactivation of TP53 and MIR34A promotes colorectal cancer development and progression in mice via increasing levels of IL6R and PAI1. Gastroenterology, 2018; 155(6): 1868-1882. doi: 10.1053/j.gastro.2018.08.011 [11] Bergeron S, Lemieux E, Durand V, et al. The serine protease inhibitor serpinE2 is a novel target of ERK signaling involved in human colorectal tumorigenesis. Mol Cancer, 2010; 9: 271. doi: 10.1186/1476-4598-9-271 [12] Dikmen S, Wang X Z, Ortega M S, et al. Single nucleotide polymorphisms associated with thermoregulation in lactating dairy cows exposed to heat stress. J Anim Breed Genet, 2015; 132(6): 409-419. doi: 10.1111/jbg.12176 [13] Li X, Zhao D, Guo Z, et al. Overexpression of SerpinE2/protease nexin-1 contribute to pathological cardiac fibrosis via increasing collagen deposition. Sci Rep, 2016; 6: 37635. doi: 10.1038/srep37635 [14] Li C, Lv L F, Qi-Li M G, et al. Endocytosis of peptidase inhibitor serpinE2 promotes myocardial fibrosis through activating ERK1/2 and β-catenin signaling pathways. Int J Biol Sci, 2022; 18(16): 6008-6601. doi: 10.7150/ijbs.67726 [15] Dong M, Yang X, Lim S, et al. Cold exposure promotes atherosclerotic plaque growth and instability via UCP1-dependent lipolysis. Cell Metab, 2013; 18(1): 118-129. doi: 10.1016/j.cmet.2013.06.003 [16] Lin A V. Direct ELISA. Methods Mol Biol, 2015; 1318: 61-67. [17] Portes A M O, Paula A B R, Miranda D C, et al. A systematic review of the effects of cold exposure on pathological cardiac remodeling in mice. J Therm Biol, 2023; 114: 103598. doi: 10.1016/j.jtherbio.2023.103598 [18] Dutta D, Donaldson J G. Search for inhibitors of endocytosis: Intended specificity and unintended consequences. Cell Logist, 2012; 2(4): 203-208. doi: 10.4161/cl.23967 [19] Joo M K, Park J J, Chun H J. Impact of homeobox genes in gastrointestinal cancer. World J Gastroenterol, 2016; 22(37): 8247-8256. doi: 10.3748/wjg.v22.i37.8247 [20] Mathijssen I M, van Leeuwen J P, Vermeij-Keers C. Simultaneous induction of apoptosis, collagen type Ⅰ expression and mineralization in the developing coronal suture following FGF4 and FGF2 application. J Craniofac Genet Dev Biol, 2000; 20(3): 127-136. [21] Lejard V, Blais F, Guerquin M J, et al. EGR1 and EGR2 involvement in vertebrate tendon differentiation. J Biol Chem, 2011; 286(7): 58555867. [22] Lincoln J, Alfieri C M, Yutzey K E. BMP and FGF regulatory pathways control cell lineage diversification of heart valve precursor cells. Dev Biol, 2006; 292(2): 292-302. [23] Li J, Salvador A M, Li G, et al. Mir-30d regulates cardiac remodeling by intracellular and paracrine signaling. Circ Res, 2021; 128(1): e1-e23. doi: 10.1161/RES.0000000000000459 [24] Gray M O, Long C S, Kalinyak J E, et al. Angiotensin Ⅱ stimulates cardiac myocyte hypertrophy via paracrine release of TGF-beta 1 and endothelin-1 from fibroblasts. Cardiovasc Res, 1998; 40(2): 352-363. doi: 10.1016/S0008-6363(98)00121-7 [25] Aceto J F, Baker K M. [Sar1]angiotensin Ⅱ receptor-mediated stimulation of protein synthesis in chick heart cells. Am J Physiol, 1990; 258(3 Pt 2): H806-H813. [26] Horio T, Maki T, Kishimoto I, et al. Production and autocrine/paracrine effects of endogenous insulin-like growth factor-1 in rat cardiac fibroblasts. Regul Pept, 2005; 124(1-3): 65-72. doi: 10.1016/j.regpep.2004.06.029 [27] Kuwahara K, Saito Y, Harada M, et al. Involvement of cardiotrophin-1 in cardiac myocyte-nonmyocyte interactions during hypertrophy of rat cardiac myocytes in vitro. Circulation, 1999; 100(10): 1116-1124. doi: 10.1161/01.CIR.100.10.1116 [28] Baker J B, Low D A, Simmer R L, et al. Protease-nexin: a cellular component that links thrombin and plasminogen activator and mediates their binding to cells. Cell, 1980; 21(1): 37-45. doi: 10.1016/0092-8674(80)90112-9 [29] Vidal R, Wagner J U G, Braeuning C, et al. Transcriptional heterogeneity of fibroblasts is a hallmark of the aging heart. JCI Insight, 2019; 4(22): e131092. doi: 10.1172/jci.insight.131092 [30] Conner S D, Schmid S L. Regulated portals of entry into the cell. Nature, 2003; 422(6927): 37-44. doi: 10.1038/nature01451 [31] McMahon H T, Boucrot E. Molecular mechanism and physiological functions of clathrin-mediated endocytosis. Nat Rev Mol Cell Biol, 2011;12(8): 517-533. doi: 10.1038/nrm3151 [32] Grainger S, Hryniuk A, Lohnes D. Cdx1 and Cdx2 exhibit transcriptional specificity in the intestine. PLoS One, 2013; 8(1): e54757. doi: 10.1371/journal.pone.0054757 [33] Pilozzi E, Onelli M R, Ziparo V, et al. CDX1 expression is reduced in colorectal carcinoma and is associated with promoter hypermethylation. J Pathol, 2004; 204(3): 289-295. doi: 10.1002/path.1641 [34] Jones M F, Hara T, Francis P, et al. The CDX1-microRNA-215 axis regulates colorectal cancer stem cell differentiation. Proc Natl Acad Sci U S A, 2015; 112(13): E1550-E1558. [35] Chu M, Wang L, Wang H, et al. A novel role of CDX1 in embryonic epicardial development. PLoS One, 2014; 9(7): e103271. doi: 10.1371/journal.pone.0103271 [36] Grabner A, Amaral A P, Schramm K, et al. Activation of cardiac fibroblast growth factor receptor 4 causes left ventricular hypertrophy. Cell Metab, 2015; 22(6): 1020-1032. doi: 10.1016/j.cmet.2015.09.002 [37] Di Marco G S, Reuter S, Kentrup D, et al. Treatment of established left ventricular hypertrophy with fibroblast growth factor receptor blockade in an animal model of CKD. Nephrol Dial Transplant, 2014; 29(11): 20282035. [38] Mitchell P, Steenstrup T, Hannon K. Expression of fibroblast growth factor family during postnatal skeletal muscle hypertrophy. J Appl Physiol (1985), 1999; 86(1): 313-319. doi: 10.1152/jappl.1999.86.1.313 [39] Lee J, Sutani A, Kaneko R, et al. In vitro generation of functional murine heart organoids via FGF4 and extracellular matrix. Nat Commun, 2020; 11(1): 4283. doi: 10.1038/s41467-020-18031-5 [40] Edalat S G, Jang Y, Kim J, et al. Collagen type Ⅰ containing hybrid hydrogel enhances cardiomyocyte maturation in a 3D cardiac model. Polymers (Basel), 2019; 11(4): 687. doi: 10.3390/polym11040687 [41] Chen X Q, Chen L L, Fan L, et al. Stem cells with FGF4-bFGF fused gene enhances the expression of bFGF and improves myocardial repair in rats. Biochem Biophys Res Commun, 2014; 447(1): 145-151. doi: 10.1016/j.bbrc.2014.03.131 -
 fzm-5-3-180_ESM.docx
fzm-5-3-180_ESM.docx

-
 点击查看大图
点击查看大图
计量
- 文章访问数: 352
- HTML全文浏览量: 191
- PDF下载量: 11
- 被引次数: 0

